
Freshwater mussels are bivalve molluscs that inhabit the beds of rivers, streams and lakes. They are found world-wide but have by far their greatest abundance and diversity in eastern North America, where over 200 native species are known. This abundance is attested to by the huge quantities of shell which were harvested by pearl-seekers and shell-button manufacturers during the late 19th and early 20th centuries (Claassen 1994) and by the enormous shell heaps that were accumulated by prehistoric humans, especially along the streams and rivers of the southeastern United States. These shell heaps or middens are generally assumed to represent the remains of shellfish gathered for consumption (e.g., Parmalee and Bogan 1986:35; Parmalee and Klippel 1974), although other explanations for their creation in particular situations have sometimes been offered (e.g., Claassen 1991). They have attracted considerable archaeological attention due to their relatively high visibility and the fact that calcareous shell tends to preserve artefacts of bone and other perishable materials.
 Left: Archaeological and modern specimens of
Elliptio crassidens, Tritagonia verrucosa, and Obliquaria
reflexa, aka the "elephant ear", "pistol grip", and "three-horned warty back".
Right: Archaeological specimens of freshwater
mussel.
Left: Archaeological and modern specimens of
Elliptio crassidens, Tritagonia verrucosa, and Obliquaria
reflexa, aka the "elephant ear", "pistol grip", and "three-horned warty back".
Right: Archaeological specimens of freshwater
mussel.
Despite the number of such sites that have been excavated, and despite the large amount of shell that has consequently been retrieved from archaeological contexts, research on the shell remains themselves has traditionally been limited to a few avenues of inquiry. Primary among these are environmental reconstruction, dietary reconstruction, and studies of seasonality. While such studies are certainly of value, there is no need for archaeologists to so limit themselves in the types of questions that can be addressed using shellfish remains. This paper will offer various suggestions for alternate research topics, along with pertinent methodological and analytical considerations. It will focus primarily on the remains of freshwater bivalves but will have implications for research on other molluscan taxa as well. It will also focus on the eastern United States, where the most abundant and diverse freshwater molluscan fauna in the world is found (Bogan 1993), but the methods may be applicable wherever extensive archaeological shell deposits exist.
It should be noted that "shell midden" is an imprecise term commonly applied to many different types of sites with varying amounts of shell in their matrices (Ambrose 1967:177; Claassen 1991). No more precise scheme for classifying shell-bearing sites has yet been adopted by archaeologists, and for the sake of simplicity "shell midden" will be used in this paper in the customary sense; that is, it will refer to any archaeological site with an appreciable amount of shell in the matrix. The utility of the methods described below will depend upon the amount, condition, and depositional context of the shell at any particular site, as well as on the cultural factors leading to the deposition of the shell. Successful application of the methods will necessitate a case-by-case evaluation of each shell assemblage to be analysed and the contexts from which the shell is retrieved.
It has frequently been assumed that prehistoric Native Americans had little effect on natural environments; indeed, North American Indians have often been characterised as 'natural conservationists' whose cultural templates precluded actions that might be considered harmful to the ecosystems within which they lived (e.g., Callicott 1982). This is an unwarranted assumption that ignores the extensive spatial, temporal and cultural variability of Native American societies as revealed through ethnology and archaeology. Zoologists and other scientists studying bivalve assemblages have been among the worst purveyors of the "pristine myth" (Denevan 1992). For example, Taylor and Spurlock describe the "pristine Ohio [River] that flowed for eons of time prior to the coming of European man" and state that "Native Americans, in apparently large numbers, had lived in harmony with the river utilizing its resources in moderation and adding nothing which would seriously harm or modify the river" (1982:49). Similarly, Taylor (1989:188) described the Ohio prior to the 18th century as a "clean, undisturbed, virgin stream" and the river valley itself as "pristine lands visited only occasionally by native-born Americans." Such statements reveal a telling ignorance of archaeological data and their implications for understanding prehistoric environmental impact. The Ohio River valley is, in fact, home to one of the richest archaeological records in the country, with good evidence of sedentary habitation and intensive land use stretching back thousands of years (Muller 1986). Rather than naive characterisations, what are needed are careful studies designed to determine the scale and effects of that land use over time.
A series of publications on the Little Tennessee River Valley, in eastern Tennessee, have shown that prehistoric land use could have a profound influence on vegetation and soils in the temperate woodlands of eastern North America (Chapman and Shea 1981; Chapman et al. 1982; H. Delcourt 1987; P. Delcourt et al. 1986). Data from a number of sources, including pollen, lacustrine stratigraphy, and charred seeds and wood fragments have shown that the landscape became progressively modified from the Late Archaic period (ca. 5,000 - 2,500 BP) onward, with significant rises in the proportions of disturbed-ground taxa at the beginnings of the succeeding Woodland and Mississippian cultural periods, during which agriculture became widespread. Paul Delcourt et al. (1986:348) have estimated that "from at least Woodland times through the Historic period, several of the lower terraces at distances of at least 1.5 km from the active floodplain were cleared of forests, placed in cultivation, or in the early states of plant succession." A concomitant increase in erosional sediment being transported into the river can logically be assumed; such an increase, if maintained over centuries, could have had a detrimental effect on the aquatic ecosystem. Freshwater mussels are particularly sensitive to siltation and are usually the first element of a stream fauna to be affected by erosional inputs (e.g., Aldridge et al. 1987; Bogan 1993:603; Cordone and Kelley 1961; Dineen 1971:193-194; Ellis 1936; Fuller 1971; Isom 1969:414; Neves 1993:3-4; Parmalee 1956:191; Parmalee et al. 1982:75; Scruggs 1960:28; Stansbery 1970:10). If prehistoric land clearance, agriculture, and other activities were being carried out at a sufficient scale, then siltation may have constituted a selective pressure that favoured more silt-tolerant mussel species over time.
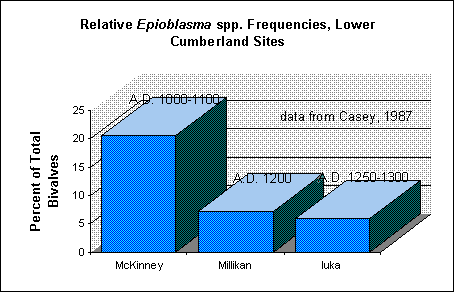 Figure 1. Relative proportions of Epioblasma spp. from three prehistoric
sites on the lower Cumberland River (data from Casey 1987).
Figure 1. Relative proportions of Epioblasma spp. from three prehistoric
sites on the lower Cumberland River (data from Casey 1987).
The molluscan data from the Little Tennessee River valley have yet to be examined in this light; however, preliminary indications from other river valleys are that the relative proportions of sensitive mussel taxa did indeed change over time. For example, the data published by Casey (1987) show a steady decrease through time in the relative proportions of Epioblasma spp. from three Mississippian sites on the lower Cumberland River, eastern Kentucky (Figure 1). Epioblasma is a genus which appears to be particularly sensitive to siltation (e.g., Dennis 1987; Neves 1993; Parmalee et al. 1980:100-101; Stansbery 1971:8). A decline in Epioblasma frequencies also can be seen in Parmalee's (1994) data from Smith Bottom Cave on the Tennessee River, from Archaic to Woodland contexts (Figure 2), and in the data
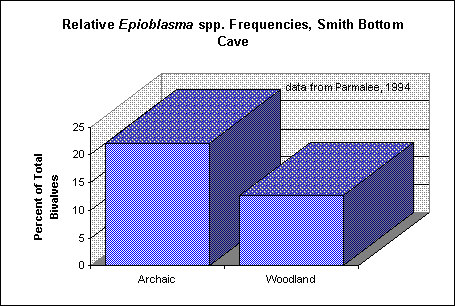 Figure 2. Relative proportions of Epioblasma spp. from Archaic (ca. 9000
- 2500 B.P.) and Woodland (ca. 2500 - 1000 B.P.) contexts in Smith Bottom Cave on the
middle Tennessee River (data from Parmalee 1994).
Figure 2. Relative proportions of Epioblasma spp. from Archaic (ca. 9000
- 2500 B.P.) and Woodland (ca. 2500 - 1000 B.P.) contexts in Smith Bottom Cave on the
middle Tennessee River (data from Parmalee 1994).presented by Parmalee et al. (1982) from Woodland to Mississippian contexts on the upper Tennessee River (Parmalee et al. 1982:86 - see Figure 3). There are a host of other factors which might explain such changes, including sampling error, the prehistoric exploitation of different stretches of the river, changes in prehistoric collection methods over time, environmental fluctuations, and so on (Casey 1987:118; Parmalee et al. 1980:94; Robison 1983:118). Still, the possibility that such changes might be attributable to erosion and sedimentation resulting from prehistoric land use must at least be considered (cf. Ambrose 1967:180). Research into this question is currently being undertaken by the author as part of doctoral research at the University of Sheffield.
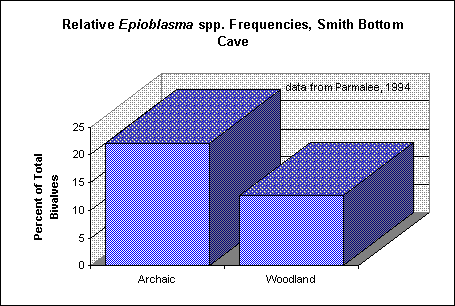
Figure 3. Relative proportions of Epioblasma spp. from Middle Woodland (ca. 2000 - 1300 B.P.), Late Woodland (ca. 1300 - 1000 B.P.) and Mississippian (ca. 1000 - 500 B.P.) sites on the Chickamauga Reservoir, upper Tennessee River (data from Parmalee et al. 1982).
Another potential use of bivalve data lies in the investigation of features as primary deposits. It is often assumed that features such as pits and postholes represent single depositional events, with the artefacts recovered from the features being treated as synchronic assemblages that ideally can be related directly to feature function. In fact, as most archaeologists are implicitly aware and as some have explicitly pointed out (Moeller 1992), features often have a mixed depositional history, especially at multicomponent sites where features originating in one stratum often intrude into one or more lower strata, thus bringing materials of various ages to the surface. These materials may then be incorporated into other features, at the end of their use (e.g., when a pit is used for trash disposal). The effect is that what is apparently a short duration phenomenon - a discrete pit - contains a long duration fill. Decoding such depositional complexities is one of the main problems facing archaeologists interested in carrying out intrasite analyses (Moeller 1992).
The presence of bivalve remains at a site may provide a means of addressing this problem. In terms of faunal analysis the quantification of bivalves is quite simple, as they have only two elements: the left and right valve. If size is also taken into account - that is, if individual specimens are separated into juvenile and adult categories - then valve representation may be used as a measure of the depositional integrity of features. Put simply, the more 'paired' valves there are, the more likely that the deposition of those valves was a single event (cf. Theler 1991:319) and that the artefacts contained within the feature therefore represent a short duration assemblage. This can be tested by seriation, a method of ordering artefact assemblages based upon stylistic change in selected artefact classes. Assemblages of non-comparable duration cannot be seriated together, as long duration assemblages will contain more types (of ceramics, or projectile points, for example) than short duration assemblages (Dunnell 1970). The bivalve data can be compared to those features identified as short duration via seriation to see if lesser post-depositional alteration of the shell assemblage can serve as a proxy for identifying short duration assemblages. If so, then the method could be used to identify short duration assemblages even in the absence of chronologically sensitive artefact types. Developing this method will require careful consideration of preservation and recovery biases; however, if successful, it could be applied to shell-bearing sites of different ages and functions world-wide.
Research undertaken within the last several years has begun to show intra- and intersite variation in the distribution of artefacts and biotic remains that has been related to social status. Scott, for example, has found that faunal assemblages at Mississippian mound centres are disproportionately weighted towards meatier elements (Scott 1982) while farmstead assemblages are weighted in the opposite direction. Blitz (1993) has shown that, at one Mississippian centre, vessels from mound contexts are larger and more open than those from non-mound contexts. These phenomena can be explained as representing either the detritus of rich provisions and vessels supplied to an elite class by an outlying population or as the remains left by communal feasting carried out at the mounds (Blitz 1993).
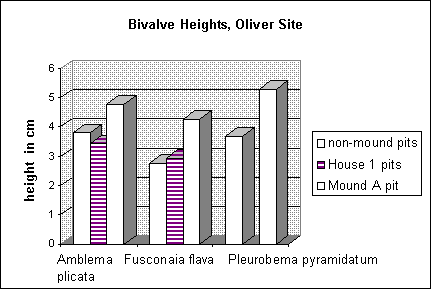
Figure 4. Average heights of selected freshwater mussel species from mound and non-mound contexts at the Oliver site, Coahoma County, Mississippi.
It may be that freshwater mussels represent another class of archaeological remains that can be used to explore this topic. Analysis of this group of molluscs from the Oliver site (22Co503), a Mississippian mound site located on the Big Sunflower River in Coahoma County, Mississippi, has revealed significant differences in the sizes of specimens retrieved from mound and non-mound contexts (Peacock n.d.). Mussels from non-mound pits were compared to those from a pit that occurred approximately in the east/southeast quadrant of Mound A, a large pyramidal mound (Duff 1994:Figure 11). Although samples were very small (Peacock n.d.), it can be seen in Figure 4 that specimens of Amblema plicata, Fusconaia flava and Pleurobema pyramidatum retrieved from the mound were much larger on average than specimens recovered from non-mound contexts. Due to the small sample sizes and the highly disturbed nature of the site (Peacock n.d.), these data must be viewed with caution. Still, it can at least be suggested that the largest mussels available were being consumed and/or discarded at the mound, in contrast to what was happening at the rest of the site. Mussels should be considered along with other biotic and artefactual remains in future explorations of these interesting patterns emerging from Mississippian sites in the region.
One of the questions that remains unanswered where prehistoric shellfish harvesting is concerned has to do with the manner in which the animals were collected. There is a dearth of ethnographic or early historic references to the topic in the eastern United States; ethnographic analogies are thus typically based upon the actions of shellfishers in Oceania or other distant areas (Claassen 1991). Most researchers seem to feel that mussels and gastropods were simply gathered by hand (e.g., Casey 1987:121; Matteson 1959:54; Robison 1983:117), although no real evidence has been presented for such an assumption. It is certainly a plausible hypothesis, given that large numbers of mussels could have been hand-collected into baskets or canoes in a short amount of time (e.g., Theler 1991). There are, however, other ways in which mussels can be collected. Mussels typically burrow partially into the substrate of a stream and open their shells to filter water and particulate matter. If a foreign object should intrude into the shell, the mussel immediately closes in a defensive reflex. This characteristic has been exploited by modern shellfish harvesters, who drag wire hooks and other implements through mussel beds: the mussels clamp down on the hooks and are then lifted from the stream bed and into the boat. If prehistoric Native Americans were aware of this behavioural trait, they could have gathered mussels simply by raking the stream bottom with the branches of trees, wooden rakes, or other implements that would not usually be preserved in the archaeological record.
There is a way in which collection methods might be explored using archaeological shell remains. Research on modern mussel assemblages has shown that healthy populations have a typical size distribution, with relatively few young individuals and a proportionately greater number of older specimens. When shell-length measurement bars are arranged upon a central axis, the distribution curve resembles an 'inverted tear drop' (Miller and Payne 1993:Figure 4; cf. Klippel et al. 1978:Figure 4; Payne and Miller 1989:Figure 1; Stober 1972:Figure 2; Strayer et al. 1981:437). If prehistoric collection methods were biased towards a particular size class - for example, if the smallest mussels were being missed, as might be expected if hand-collecting was the method being employed (Murphy 1971:22; Parmalee et al. 1982:81; Parmalee and Bogan 1986:35; Robison 1983:117) - then this should be reflected in size measurements taken on archaeological specimens (cf. Klippel et al. 1978; Parmalee 1956:189). A cursory examination of shell from the Oliver site (Peacock n.d.) showed that mussels of all sizes, including very small ones, were being taken, a situation that has been noted elsewhere in the region (e.g., Parmalee and Bogan 1986:29-30, 35). This fact, coupled with the common presence of small aquatic gastropods in shell middens (e.g., Parmalee and Bogan 1986:35), suggests that some form of harvesting other than hand-collecting might have been employed in the past. Size analysis, coupled with experimental archaeology, should help to shed some light on prehistoric shellfish collection methods in the future.
Bivalve analysis in archaeology has come a long way since Morse (1925:429) described coastal middens as "the refuse piles or kitchen refuse of a rude and savage people." Environmental reconstruction, dietary reconstruction, and studies of seasonality have all contributed to our understanding of prehistoric societies. However, as this paper has attempted to show, there are numerous other types of questions that can addressed using shellfish remains, including the investigation of anthropogenic impact, contextual concerns, social differences in resource distribution, and prehistoric shellfishing methods.
In order to address such questions, it is imperative that archaeologists treat shellfish remains as they would any other type of artefact: that is, such remains must be systematically collected, processed and curated. All too often shellfish remains are 'sampled' in some fashion and the bulk discarded. Such sampling, even if done in a systematic fashion, greatly diminishes the chances of addressing questions such as those outlined above. In the study of anthropogenic impact, for example, large numbers of specimens are needed in order to characterise past mussel communities (Miller and Payne 1988, 1993), and it should be remembered that the rarer specimens - those most likely to be lost when sampling is employed - are the most sensitive indicators of past environmental conditions. The investigation of intrasite differences in mussel species or size distributions (e.g., Theler 1991:323) likewise necessitates the retention of as much shell as possible. Archaeologists have a responsibility to their discipline, as well as to zoologists, biogeographers, ecologists and other scientists (cf. Barber 1982; Bogan 1981:33; Murphy 1971), to curate all the shell remains recovered from archaeological sites. To do less is to diminish the amount that we can learn about the past societies that produced the middens, and the world of which those societies were a part.
Aldridge, D.W., B.S. Payne, and A.C. Miller. 1987. The effects of intermittent exposure to suspended solids and turbulence on three species of freshwater mussel. Environmental Pollution 45:17-28.
Ambrose, W.R. 1967. Archaeology and shell middens. Archaeology and Physical Anthropology in Oceania 2(3):169-187.
Barber, R.J. 1982. The indigenous distribution of Elliptio complanata in Ohio: archaeological evidence. The Nautilus 96(4):130-131.
Blitz, J.H. 1993. Big pots for big shots: feasting and storage in a Mississippian community. American Antiquity 58(1):80-96.
Bogan, A.E. 1981. A reconstruction of the freshwater molluscan fauna of the Little Tennessee River, east Tennessee. The Bulletin of the American Malacological Union for 1981:33-34.
Bogan, A.E. 1993. Freshwater bivalve extinctions (Mollusca: Unionoida): a search for causes. American Zoologist 33:599-609.
Callicott, J.B. 1982. Traditional American Indian and western European attitudes toward nature: an overview. Environmental Ethics 4:293-318.
Casey, J.L. 1987. Aboriginal and modern mussel assemblages of the lower Cumberland River. Southeastern Archaeology 6(2):115-124.
Chapman, J., and A.B. Shea. 1981. The archaeobotanical record: Early Archaic period to Contact in the lower Little Tennessee River Valley. Tennessee Anthropologist 6(1):61-84.
Chapman, J., P.A. Delcourt, P.A. Cridlebaugh, A.B. Shea, and H.R. Delcourt. 1982. Man-land interactions: 10,000 years of American Indian impact on native ecosystems in the lower Little Tennessee River Valley, eastern Tennessee. Southeastern Archaeology 1(2):115-121.
Claassen, C. 1991. Normative thinking and shell-bearing sites. In M.B. Schiffer (ed.), Archaeological Method and Theory 3:249-298. Tuscon: The University of Arizona Press.
Claassen, C. 1994. Washboards, pigtoes and muckets: historic musseling in the Mississippi watershed. Historical Archaeology 28(2).
Cordone, A.J., and D.W. Kelley. 1961. The influence of inorganic sediment on the aquatic life of streams. California Fish and Game 47:189-228.
Delcourt, H.R. 1987. The impact of prehistoric agriculture and land occupation on natural vegetation. Trends in Ecology and Evolution 2(2):39-44..
Delcourt, P.A., H.R. Delcourt, P.A. Cridlebaugh, and J. Chapman. 1986. Holocene ethnobotanical and paleoecological record of human impact on vegetation in the Little Tennessee River Valley, Tennessee. Quaternary Research 25:330-349.
Denevan, W.M. 1992. The Pristine Myth: the landscape of the Americas in 1492. Annals of the Association of American Geographers 82:369-385.
Dennis, S.D. 1987. An unexpected decline in populations of the freshwater mussel Dysnomia (= Epioblasma) capsaeformis in the Clinch River of Virginia and Tennessee. Virginia Journal of Science 38(4):281-288.
Dineen, C.F. 1971. Changes in the molluscan fauna of the Saint Joseph River, Indiana, between 1959 and 1970. Transactions of the Indiana Academy of Science 80:189-195.
Duff, E.A. 1994. Recent Salvage Excavations at the Oliver Site and an Evaluation of Mississippian Period Chronology in the Upper Yazoo Basin of Mississippi. Thesis submitted to the Department of Sociology and Anthropology, University of Mississippi.
Dunnell, R.C. 1970. Seriation method and its evaluation. American Antiquity 35:305-319.
Ellis, M.M. 1936. Erosion silt as a factor in aquatic environments. Ecology 17(1):29-42.
Fuller, S.L.H. 1971. A brief guide to the fresh-water mussels (Mollusca: Bivalvia: Unionacea) of the Savannah River system. ASB Bulletin 18(4):139-146.
Isom, B.G. 1969. The mussel resource of the Tennessee River. Malacologia 7:397-425.
Klippel, W.E., G. Celmer, and J.R. Purdue. 1978. The Holocene naiad record at Rodgers Shelter in the western Ozark Highlands of Missouri. Plains Anthropologist 23 (No. 82, Part 1):257-271.
Matteson, M.R. 1959. An analysis of the shells of fresh-water mussels gathered by Indians in southwestern Illinois. Transactions of the Illinois Academy of Science 52:52-58.
Miller, A.C., and B.S. Payne. 1988. The need for quantitative sampling to characterise size demography and density of freshwater mussel communities. American Malacological Bulletin 6(1):49-54.
Miller, A.C. and B.S. Payne. 1993. Qualitative versus quantitative sampling to evaluate population and community characteristics at a large-river mussel bed. American Midland Naturalist 130:133-145.
Moeller, R.W. 1992. Analyzing and interpreting Late Woodland features. Occasional Publications in Northeastern Anthropology No. 12.
Morse, E.S. 1925. Shell-mounds and changes in the shells composing them. The Scientific Monthly 21:429-440.
Muller, J. 1986. Archaeology of the Lower Ohio River Valley. Orlando: Academic Press.
Murphy, J.L. 1971. Molluscan remains from four archaeological sites in northeastern Ohio. Sterkiana 43:21-25.
Neves, R.J. 1993. A state-of-the-unionids address. In K.S. Cummings, A.C. Buchanan, and L.M. Koch (eds.), Conservation and Management of Freshwater Mussels, pp. 1-10. Rock Island, Illinois: Upper Mississippi River Conservation Committee.
Parmalee, P.W. 1956. A comparison of past and present populations of fresh-water mussels in southern Illinois. Illinois Academy of Science Transactions 49:184-192.
Parmalee, P.W. 1994. Freshwater mussels from Dust and Smith Bottom Caves, Alabama. In N.S. Goldman-Finn and B.N. Driskell (eds.), Preliminary Archaeological Papers on Dust Cave, Northwest Alabama. Journal of Alabama Archaeology 40 (1 and 2):135-162.
Parmalee, P.W., and A.E. Bogan. 1986. Molluscan remains from aboriginal middens at the Clinch River Breeder Reactor Plant Site, Roane County, Tennessee. American Malacological Bulletin 4(1):25-37.
Parmalee, P.W., and W.E. Klippel. 1974. Freshwater mussels as a prehistoric food resource. American Antiquity 39(3):421-434.
Parmalee, P.W., W.E. Klippel, and A.E. Bogan. 1980. Notes on the prehistoric and present status of the naiad fauna of the middle Cumberland River, Smith County, Tennessee. The Nautilus 94(3):93-105.
Parmalee, P.W., W.E. Klippel, and A.E. Bogan. 1982. Aboriginal and modern freshwater mussel assemblages (Pelecypoda: Unionidae) from the Chickamauga Reservoir, Tennessee. Brimleyana 8:75-90.
Payne, B.S., and A.C. Miller. 1989. Growth and survival of recent recruits to a population of Fusconaia ebena (Bivalvia: Unionidae) in the lower Ohio River. American Midland Naturalist 121:99-104.
Peacock, E. n.d. Mussel Shell and Gastropod Analysis from the 1990-1991 Salvage. In J. M. Connaway, J. K. Johnson, and P. Galloway (eds.), The Oliver Site (22-Co-503), Coahoma County, Mississippi: Collected Papers. Ms. in preparation, to be published by the Mississippi Department of Archives and History, Jackson, Mississippi, U.S.A.
Robison, N.D. 1983. Archeological records of naiad mussels along the Tennessee-Tombigbee Waterway. In A. C. Miller (compiler), Report of Freshwater Mussels Workshop, 26-27 October 1982, pp. 115-129. Vicksburg, Mississippi: U. S. Army Corps of Engineers, Waterways Experiment Station, Environmental Laboratory.
Scott, S. 1982. Yarborough Site faunal remains. In C. Solis and R. Walling, Archaeological Investigations at the Yarborough Site (22Cl814), Clay County, Mississippi. University of Alabama, Office of Archaeological Research, Report of Investigations 30, pp. 140-152.
Scruggs, G.D., Jr. 1960. Status of fresh-water mussel stocks in the Tennessee River. U.S. Fish and Wildlife Service Special Scientific Report - Fisheries No. 370.
Stansbery, D.H. 1970. American Malacological Union Symposium, Rare and Endangered Mollusks. 2. Eastern Freshwater Mollusks (I) The Mississippi and St. Lawrence River systems. Malacologia 10:9-22.
Stansbery, D.H. 1971. Rare and endangered freshwater mollusks in eastern United States. In S.E. Jorgensen and R.W. Sharp (eds.), Proceedings of a Symposium on Rare and Endangered Mollusks (Naiads) of the U. S., pp. 5-18. U. S. Dept. of the Interior, Fish and Wildlife Service, Bureau of Sport Fisheries and Wildlife.
Stober, Q.J. 1972. Distribution and age of Margaritifera margaritifera (L.) in a Madison River (Montana, U.S.A.) mussel bed. Malacologia 11:343-350.
Strayer, D.L., J.J. Cole, G.E. Likens, and D.C. Buso. 1981. Biomass and annual production of the freshwater mussel Elliptio complanata in an oligotrophic softwater lake. Freshwater Biology 11:435-440.
Taylor, R.W. 1989. Changes in freshwater mussel populations of the Ohio River: 1,000 BP to recent times. Ohio Journal of Science 89(5):188-191.
Taylor, R.W., and B.D. Spurlock. 1982. The changing Ohio River naiad fauna: a comparison of early Indian middens with today. The Nautilus 96(2):49-51.
Theler, J.L. 1991. Aboriginal utilization of freshwater mussels at the Aztalan Site, Wisconsin. In J.R. Purdue, W.E. Klippel, and B.W. Styles (eds.,), Beamers, Bobwhites, and Blue-Points: Tributes to the Career of Paul W. Parmalee, pp. 315-332. University of Tennessee, Department of Anthropology, Report of Investigations 52.
About the author: Evan Peacock received a B.A. in anthropology from Mississippi State University in 1988 and a M.Sc. in Environmental Archaeology and Palaeoeconomy from the University of Sheffield in 1990. He is currently pursuing a Ph.D. at Sheffield on research being carried out at prehistoric shell middens in the southeastern United States. Until his thesis is finished, he is not allowed to have any outside interests.
�Evan Peacock 1996

�assemblage 1996