On The Evolution of Human Aesthetic Preferences.*
Andrew T. Chamberlain
How can we imagine that an inch in the tail of the peacock, or ¼
inch in that of the Bird of Paradise, would be noticed and preferred by
the female? Alfred Wallace, letter to Charles Darwin, March 1868.
In regard to sexual selection. A girl sees a handsome man, and without
observing whether his nose or whiskers are the tenth of an inch longer
or shorter than in some other man, admires his appearance and says she
will marry him. So, I suppose, with the pea-hen ..... Charles Darwin,
letter to Alfred Wallace, March 1868.
Introduction
The archaeological record of the last Ice Age, particularly after about
40,000 years ago, documents the emergence of clearly symbolic behaviour,
including burials that were furnished with grave goods, the invention of
parietal and portable representational art, the use of items of body ornamentation
and possible instances of numerical or calendrical notation (Pfeiffer,
1982). This kind of material cultural evidence has informed and stimulated
the discussion of the evolution of modern human cognitive abilities, but
there is also an important aesthetic dimension to human culture, and as
Kaplan (1992) has emphasised, evolutionary explanations
of human aesthetic preferences benefit from integrated approaches that
consider both cognitive and emotional responses. On both theoretical and
empirical grounds it is likely that human emotional adaptations evolved
much earlier than strictly symbolic capacities, given that 99% of the five
million year timespan of hominid evolution preceded the emergence of material
evidence for symbolic behaviour. Thus the study of human aesthetic preferences
may provide a window into an extensive epoch in the early evolution of
the human psyche.
In this paper I review some human visual aesthetic preferences that
may have originated in our species distant evolutionary past. These preferences
include evolved responses to natural landscapes, symmetry preferences,
and criteria of facial attractiveness. In the discussion that follows I
employ a very general definition of aesthetics, best summarised as "mental
appreciation of the shape or embellishment imposed on raw materials" (cf.
Dissanayake, 1992), in which the term "appreciation" primarily denotes
an involuntary emotional response to a stimulus, rather than the deliberate
intellectual stance adopted by the modern professional or philosophical
aesthete.
Landscape Preferences and the Hominid Environment of Evolutionary Adaptiveness
 An
innate human preference for visual landscapes that have properties resembling
those of savanna habitats (i.e. low-relief, sparsely-wooded tropical grasslands:
Figure 1a & b) has been attributed to selection pressures
operating during early human evolution (Balling &
Falk, 1982; Orians & Heerwagen, 1992).
According to Orians and Heerwagen, present-day humans express a rapid and
often unconscious affective response to those general properties of a landscape
that are perceived on initial visual encounter. Preferred landscapes are
those containing features indicative of environmental conditions favourable
for survival, such as an abundance of subsistence resources or a minimal
threat from predators. The preference for savanna-style visual landscapes
is most strongly expressed in children (Balling &
Falk, 1982), and is also manifest in the deliberate design of artificial
landscapes as exemplified by modern (i.e. post-Renaissance) ornamental
parks and gardens (Kaplan, 1992). A more specific
hypothesis of landscape preference stems from "prospect-refuge theory",
which predicts that within a given landscape preferred locations are found
at interfaces between prospect-dominant and refuge-dominant areas (Appleton,
1996). These vantage points combine unimpeded visual prospects with
a ready opportunity for concealment and/or withdrawal to a safe refuge.
Thus a treeless landscape is less visually attractive than a habitat containing
isolated trees that can provide opportunities to hide or escape from potential
predators.
An
innate human preference for visual landscapes that have properties resembling
those of savanna habitats (i.e. low-relief, sparsely-wooded tropical grasslands:
Figure 1a & b) has been attributed to selection pressures
operating during early human evolution (Balling &
Falk, 1982; Orians & Heerwagen, 1992).
According to Orians and Heerwagen, present-day humans express a rapid and
often unconscious affective response to those general properties of a landscape
that are perceived on initial visual encounter. Preferred landscapes are
those containing features indicative of environmental conditions favourable
for survival, such as an abundance of subsistence resources or a minimal
threat from predators. The preference for savanna-style visual landscapes
is most strongly expressed in children (Balling &
Falk, 1982), and is also manifest in the deliberate design of artificial
landscapes as exemplified by modern (i.e. post-Renaissance) ornamental
parks and gardens (Kaplan, 1992). A more specific
hypothesis of landscape preference stems from "prospect-refuge theory",
which predicts that within a given landscape preferred locations are found
at interfaces between prospect-dominant and refuge-dominant areas (Appleton,
1996). These vantage points combine unimpeded visual prospects with
a ready opportunity for concealment and/or withdrawal to a safe refuge.
Thus a treeless landscape is less visually attractive than a habitat containing
isolated trees that can provide opportunities to hide or escape from potential
predators.
 Exponents
of the human preference for savanna-like habitats have reasoned that the
human environment of evolutionary adaptiveness (EEA) was located in the
Plio-Pleistocene savannas of sub-Saharan Africa. It has been argued that
habitual occupation of the savanna biome by Australopithecus and
early species of Homo provided an extended period of selection for
the reinforcement of intuitive preferences for certain topographic, botanical
and faunal features of the savanna landscape (cf. Orians
& Heerwagen, 1992: 556). However, this scenario of human evolution
is over-simplified, and there is an increasing consensus among palaeoanthropologists
that there is no single unitary environment to which earlier human species
were optimally adapted (Foley, 1996).
Ardipithecus
ramidus and Australopithecus anamensis, the earliest known hominids,
show morphological adaptations to arboreal substrates (White
et al, 1994; Leakey et al., 1998) and the fossils
of Ardipithecus ramidus have been recovered from depositional contexts
characteristic of closed canopy woodland rather than open savanna (WoldeGabriel
et al., 1994). Furthermore there is also an extensive and more recent
history of human occupation of non-savanna biomes, especially after 1.8
million years ago when species of Homo first appear in higher latitude
regions of Asia and Europe.
Exponents
of the human preference for savanna-like habitats have reasoned that the
human environment of evolutionary adaptiveness (EEA) was located in the
Plio-Pleistocene savannas of sub-Saharan Africa. It has been argued that
habitual occupation of the savanna biome by Australopithecus and
early species of Homo provided an extended period of selection for
the reinforcement of intuitive preferences for certain topographic, botanical
and faunal features of the savanna landscape (cf. Orians
& Heerwagen, 1992: 556). However, this scenario of human evolution
is over-simplified, and there is an increasing consensus among palaeoanthropologists
that there is no single unitary environment to which earlier human species
were optimally adapted (Foley, 1996).
Ardipithecus
ramidus and Australopithecus anamensis, the earliest known hominids,
show morphological adaptations to arboreal substrates (White
et al, 1994; Leakey et al., 1998) and the fossils
of Ardipithecus ramidus have been recovered from depositional contexts
characteristic of closed canopy woodland rather than open savanna (WoldeGabriel
et al., 1994). Furthermore there is also an extensive and more recent
history of human occupation of non-savanna biomes, especially after 1.8
million years ago when species of Homo first appear in higher latitude
regions of Asia and Europe.
Evidence of heterogeneity in hominid landscape preferences has been
sought in studies of the geological and ecological contexts within which
hominid fossils have been discovered, in an attempt to delineate ecological
differences between hominid species (Behrensmeyer,
1978; White, 1988). There is a substantial and
difficult taphonomic problem here: are the habitats that are conducive
to the deposition and preservation of hominid fossils representative of
the environments to which the hominids were originally and optimally adapted?
Whites (1988) analysis of the relative frequencies
of Homo and Australopithecus fossils in different African
palaeoenvironments failed to confirm earlier suggestions that Homo
fossils were relatively more common in river bank and lake margin settings.
However, most of the East and South African Pleistocene hominid sites have
palaeoenvironmental profiles indicating a mosaic habitat, with a mixture
of flora and fauna that are individually adapted to open grassland, woodland
and proximity to water. This provides indirect support for the prospect-refuge
theory of human landscape preference, as the palaeoenvironmental evidence
would be expected to show a mixture of habitat types if hominid occupation
sites were preferentially located at habitat interfaces.
Thus if there is an evolved human psychological preference for savanna
habitats this is unlikely to have been inherited as a phylogenetic legacy
from a much earlier (i.e. australopithecine) phase of human evolution.
The palaeoecological evidence, though sparse, suggests that Lower Pleistocene
hominids favoured either closed woodland (Ardipithecus) or mosaic
habitats (Australopithecus and early Homo). Furthermore,
in living primates there are instances of divergence of substrate and habitat
preference between closely-related species (for instance, both arboreal
and terrestrial species exist within each of the genera Cercopithecus
and Macaca), suggesting that evolved environmental preferences in
primates are typically species-specific adaptations rather than plesiomorphic
traits inherited by all members of a genus. This raises the possibility
that the savanna landscape preference that characterises modern humans
arose relatively recently, perhaps coinciding with the emergence of anatomically
modern Homo sapiens. One model of the origin of anatomically modern
humans places the crucial speciation event that gave rise to our species
in sub-Saharan Africa, at about 100,000 years ago (Stringer
& Andrews, 1988). It is therefore possible that the preference
for savanna habitats emerged when modern humans were confined to these
regions of Africa, prior to their expansion to other regions of the world
during the Upper Pleistocene.
Symmetry Preferences
Geometrical symmetry is widespread in nature and in human culture. A
feature of both the natural and cultural realms is that symmetry of form
is frequently over-determined in relation to functional necessity: that
is, organisms and artefacts often appear to be more symmetrical than is
strictly necessary for the purposes of efficient mechanical design. In
animals, the apparent regularity of bilateral symmetry is sometimes literally
skin-deep: dissection reveals that the thoracic and abdominal organs of
humans (and other vertebrate animals) are profoundly asymmetrical. In like
fashion, artefacts often exhibit patterns of symmetry that are not dictated
solely by technological considerations (Wynn, 1995,
2000; Kohn & Mithen, 1999).
A large body of experimental evidence supports the hypothesis that many
species of animals use symmetry in the external appearance of conspecifics
as a visual or tactile criterion in mate selection. Symmetry preferences
may emerge either as an unintentional by-product of the evolution of visual
shape recognition mechanisms (Enquist & Arak,
1994; Johnstone, 1994) or because symmetry
is an effective and easily perceived measure of genetic quality (Møller,
1990; Parsons, 1992). As both sides of a bilaterally
symmetrical animal are usually products of the same genetic code, small
deviations from symmetry between the left and right sides of an organism
serve as a proxy measure of how faithfully the animals genotype has been
expressed during its development. Individual animals differ in their degree
of random departures from symmetry because they differ in their ability
to execute developmental programs accurately when challenged by environmental
stress: symmetry is thus a sensitive indicator of the genotypes ability
to canalise development and of its resistance to environmental stress.
The morphological ideal of symmetry is easily conceptualised, and even
small deviations from the ideal are therefore detectable and measurable
by simple perceptual systems.
Visually striking external weapons and ornaments serve as effective
"honest advertisements" of an animals phenotypic quality. Single ornaments
(e.g. horns in rhinoceroses) are placed centrally, and paired display structures
(e.g. antlers in deer and tusks in elephants) are positioned with sufficient
proximity to each other to facilitate rapid visual assessment of their
degree of symmetry in size and shape. Selection of mating partners places
secondary sexual traits or ornaments under strong directional selection,
and an additional effect is that the symmetry and the size of an ornament
become positively intercorrelated, that is individuals with the largest
ornaments tend also to be more symmetrical (Møller,
1990). Thus individuals with superior quality or condition can advertise
their status by developing both large and symmetrical traits, and since
this is a strategy that inferior animals are unable to mimic these structures
serve as "honest" markers of fitness (Zahavi &
Zahavi, 1997).
In male non-human primates the enlarged upper canine teeth serve as
display organs, and the degree of symmetry between the size of the right
and left side canine teeth is correlated with the intensity of sexual selection
(Manning
& Chamberlain, 1993). In strongly sexually dimorphic primate species,
such as the western lowland gorilla, the male animals with the largest
canine teeth are also those with the most symmetrical teeth (Manning
& Chamberlain, 1994), indicating that for these animals there is
intraspecific selection for both size and symmetry in the canine teeth.
Hominid species, in contrast, have reduced canine teeth that do not project
significantly beyond the occlusal plane, yet early hominids of the genera
Ardipithecus, Australopithecus and Paranthropus show
marked sexual dimorphism in body size, an attribute which is consistent
with elevated levels of inter-male competition. Canine reduction in hominids
coincides with the appearance of bipedal posture and locomotion, and it
is possible that active bipedal displays replaced static canine displays
in agonistic encounters between early hominids (Jablonski
& Chaplin, 1993).
 Unfortunately
there are too few sufficiently well-preserved fossil specimens to test
for the possible effects of sexual selection on the symmetry of canine
teeth of early hominids, but from about 2½ million years ago an
increasingly rich archaeological record emerges providing an opportunity
to examine symmetry preferences in the manufacture of stone tools. As Wynn
(1995, 2000) has shown, simple
worked cores and flaked stone tools of the Oldowan (Mode 1) cultural repertoire
do not incorporate a principle of symmetry, but in Acheulian bifacial (Mode
2) production sophisticated symmetries in three dimensions are achieved
(Figure
2). Acheulian handaxes are clearly over-determined in their symmetry
of outline in relation to their design and use as cutting or chopping implements.
This implies that in addition to a strictly utilitarian role they may have
served as display items, perhaps as signs of the perceptual and productive
skills of their makers, or simply as aesthetically satisfying objects
(Kohn
& Mithen, 1999). This conjecture that these artefacts are display
items is testable because, as with visually prominent organic weapons and
ornaments, we would expect a positive correlation between the size and
the overall symmetry of the artefact. The correlation arises because both
large size and symmetry enhance the visual impact of a handaxe, so the
most impressive artefacts are those which excel in both attributes. As
handaxes are manufactured by reduction from a large core it is exceedingly
difficult to manufacture large and symmetrical items - symmetry is only
achieved by removal of stone - and a large symmetrical axe will
therefore serve as an 'honest' advertisement of its maker's knapping skills.
Unfortunately
there are too few sufficiently well-preserved fossil specimens to test
for the possible effects of sexual selection on the symmetry of canine
teeth of early hominids, but from about 2½ million years ago an
increasingly rich archaeological record emerges providing an opportunity
to examine symmetry preferences in the manufacture of stone tools. As Wynn
(1995, 2000) has shown, simple
worked cores and flaked stone tools of the Oldowan (Mode 1) cultural repertoire
do not incorporate a principle of symmetry, but in Acheulian bifacial (Mode
2) production sophisticated symmetries in three dimensions are achieved
(Figure
2). Acheulian handaxes are clearly over-determined in their symmetry
of outline in relation to their design and use as cutting or chopping implements.
This implies that in addition to a strictly utilitarian role they may have
served as display items, perhaps as signs of the perceptual and productive
skills of their makers, or simply as aesthetically satisfying objects
(Kohn
& Mithen, 1999). This conjecture that these artefacts are display
items is testable because, as with visually prominent organic weapons and
ornaments, we would expect a positive correlation between the size and
the overall symmetry of the artefact. The correlation arises because both
large size and symmetry enhance the visual impact of a handaxe, so the
most impressive artefacts are those which excel in both attributes. As
handaxes are manufactured by reduction from a large core it is exceedingly
difficult to manufacture large and symmetrical items - symmetry is only
achieved by removal of stone - and a large symmetrical axe will
therefore serve as an 'honest' advertisement of its maker's knapping skills.
Although I do not have access to appropriate data on the symmetry of
Lower Palaeolithic artefacts I have measured the symmetry of a large sample
of British Neolithic stone axes for which the cross-sectional shapes have
been published (Adkins & Jackson, 1978). Circumstantial
evidence suggests that these artefacts had a significant display value
- they were sometimes made on precious raw materials and are distributed
many hundreds of kilometres from the source of stone, they were frequently
polished on all surfaces and not simply on the cutting edge, and they are
often discovered in an unused condition and are sometimes clustered in
hoards and in votive deposits. The cross-sectional asymmetry of a sample
of 333 Neolithic axes recovered from the River Thames in southern England
shows a small but significant negative correlation with the length of the
artefact, as expected if both size and symmetry of outline form were preferred
criteria. Small axes may, but need not be symmetrical, but large axes are
much more likely to have a symmetrical cross section (Figure
3).
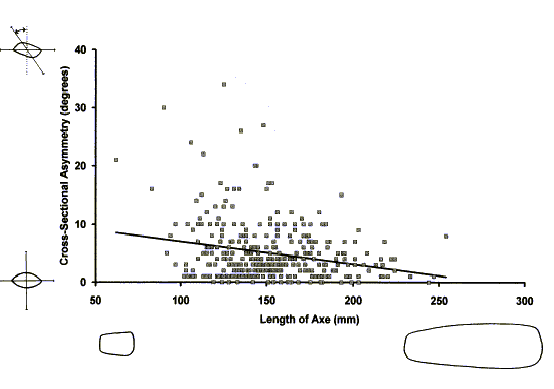 Figure
3: The relationship between cross-sectional asymmetry and length in
Neolithic stone axes. Asymmetry is measured from cross-sectional profiles
as the deviation in degrees of the axis of maximum thickness from a line
drawn at right angles to the axis of maximum breadth. The axis of maximum
thickness is defined as the line connecting the two points where lines
that are parallel to the axis of maximum breadth make tangential contact
with the cross-sectional outline of the axe. Asymmetry is negatively correlated
with the length of the axe: r = -0×24, n=233, p<< 0 × 01
Figure
3: The relationship between cross-sectional asymmetry and length in
Neolithic stone axes. Asymmetry is measured from cross-sectional profiles
as the deviation in degrees of the axis of maximum thickness from a line
drawn at right angles to the axis of maximum breadth. The axis of maximum
thickness is defined as the line connecting the two points where lines
that are parallel to the axis of maximum breadth make tangential contact
with the cross-sectional outline of the axe. Asymmetry is negatively correlated
with the length of the axe: r = -0×24, n=233, p<< 0 × 01
Mithen (1997) has argued that the archaic minds
of pre-modern hominid species had 'domain specific' intelligence in which
the modules concerned with interacting with the natural world were essentially
isolated from the mental modules employed in the manufacture and use of
artefacts. The presence of patterns of symmetry in Lower Palaeolithic artefacts
that mimic those found in biological weapons and ornaments suggests that
aesthetic preferences, at least, may be less constrained by domain specificity
than has been argued in the case of more explicitly cognitive properties
of the evolving human mind.
Facial Attractiveness
It is likely that humans and non-human primates share evolved mechanisms
that specialise in facial recognition and in the interpretation of facial
emotional expressions. In humans these mechanisms for processing visual
data from facial configurations are augmented by shared, possibly species-specific,
preferences in facial attractiveness. While non-human primates are similar
to humans in some aspects of their face recognition mechanisms (e.g. Phelps
& Roberts, 1994) it is not yet clear whether facial attractiveness
is an important criterion of mate selection in primates, although the vivid
and often sexually dimorphic facial colouration of guenons and mandrills
is believed to result from directional sexual selection (Kingdon,
1980).
A large amount of research has been published on determinants of facial
attractiveness in humans, but many studies have been restricted in scope,
for example evaluating exclusively preferences for female faces or confining
their analyses to western (Caucasian) populations. As with landscape preferences,
differential interest in attractive adult female faces emerges early in
the first year of infancy implying that the preference for attractive faces
is more likely to be an evolved response than a learned behaviour (Langlois
et al, 1987). There is also a significant degree of cross-cultural
agreement between tests of ratings of facial attractiveness conducted in
populations of different ethnic backgrounds (Jones et
al., 1993; Perrett et al., 1994). Taken together
this evidence disproves the argument that criteria of physical attractiveness
are determined by arbitrary cultural conventions.
What are the structural characteristics of human faces that influence
judgements of facial attractiveness? Artificial facial images constructed
from composites of a sample of different faces are rated as being more
attractive than the individual source images (Symons,
1979; Langlois & Roggman, 1990). This
finding of a preference for a stereotypical average has been cited as evidence
for the effect of stabilising selection on facial shape, but the computer
graphical processing of images to generate a sample average increases facial
symmetry and eliminates facial irregularities, and this "smoothing" effect
in itself makes facial composites appear more youthful (Burt
& Perrett, 1995). Males especially show heightened preferences
for younger female faces, and it is therefore difficult to ascertain whether
"averageness" per se is attractive (Alley &
Cunningham, 1991). Individual faces that are rated as highly attractive
are in fact not average but depart from the sample and population mean
in 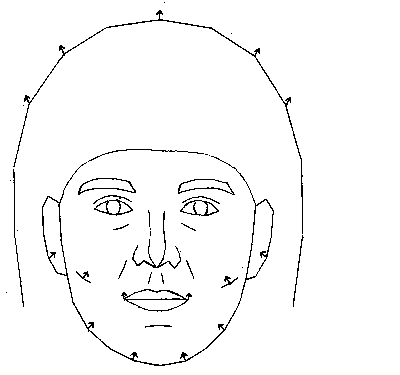 predictable
ways, and artificial exaggeration of distinctive differences from the average
face through automated caricaturisation enhances facial attractiveness
(Perrett et al., 1994). In Perrett et al.s study of Caucasian and Japanese
subjects, attractive female faces in both populations had a thinner lower
jaw, shorter lower facial height, higher forehead and relatively larger
eyes (Figure 4). These characters are neotenous in
humans and therefore contribute to a youthful facial appearance in adults
(Jones, 1996).
predictable
ways, and artificial exaggeration of distinctive differences from the average
face through automated caricaturisation enhances facial attractiveness
(Perrett et al., 1994). In Perrett et al.s study of Caucasian and Japanese
subjects, attractive female faces in both populations had a thinner lower
jaw, shorter lower facial height, higher forehead and relatively larger
eyes (Figure 4). These characters are neotenous in
humans and therefore contribute to a youthful facial appearance in adults
(Jones, 1996).
Figure 4: Outline of a composite face constructed from the average
of 60 Caucasian females (redrawn from Perrett et al.,
1994). Arrows indicate the vectors of change of topographical landmarks
when the average face is morphed onto the mean of the 15 faces with the
highest attractiveness ratings. These vectors show that the stereotypical
attractive face has a thinner lower jaw, reduced lower facial height and
a higher forehead than the population average.
Human facial preferences may have a deep evolutionary history. The morphological
characters of the female face that influence attractiveness are also those
which have been subject to directional evolutionary change during human
evolution, particularly during the Pleistocene evolution of the genus Homo,
and they are part of a suite of uniquely derived morphological characteristics
that distinguish Homo sapiens from all other hominids. It is therefore
possible that current human facial preferences are founded on a legacy
of directional selection for characteristics that are important in the
human specific-mate recognition system. This hypothesis can only be tested
indirectly on the skeletal remains of fossil hominids (faces do not fossilise,
and forensic methods of reconstructing faces from skeletonised remains
are inherently unreliable).
It is also noteworthy that neotenous facial proportions elicit protective
and nurturing behaviour in human caregivers (McCabe,
1988), and the same effect can be observed in other mammals (Lorentz,
1971). Elicitation of parental care may be of heightened importance
to humans because of the extended period during which human offspring are
dependent on the support of adult kin. Criteria of human facial attractiveness
may therefore reflect a generalised mammalian preference for neotenic facial
forms, and it is interesting to note that human standards of attractiveness
are frequently applied to animals and may be implicated in the morphological
changes that have accompanied the domestication and selective breeding
of animal species (Clutton-Brock, 1981).
Conclusion
This short review has only examined one sensory modality - vision -
and discussion has been limited to three arenas within which visual aesthetic
preferences appear to have an evolutionary origin. Evolved responses to
aspects of the visual environment appear to be a palimpsest of adaptive
strategies of greater and lesser antiquity (Table 1).
Table 1: Possible Origins and Functions of Human Aesthetic Preferences
| Preference |
Hypothesised Origin |
Postulated Function |
| Savanna Landscape |
Anatomically modern Homo sapiens |
Foraging and defence |
| Symmetry in Organisms and Artefacts |
Generalised feature of mate selection strategies in vertebrates |
Honest advertisement and mate selection |
| Facial Attractiveness |
General mammalian preference, perhaps increased in importance in
early Homo |
Mate selection, and elicitation of nurturing behaviour |
Landscape preferences are a priori likely to be species-specific,
and the "savannah" preference documented for modern humans can be interpreted
in the light of the "Out of Africa" model which posits a recent sub-Saharan
origin for our species. Symmetry preferences may have originated in mechanisms
that evolved for monitoring phenotypic quality in conspecific mates: such
mechanisms are extremely widespread in nature, and the fact that the symmetry
preference is easily transferred to items of material culture that have
semiotic value suggests that the preference may serve a domain-general
function in humans. Facial attractiveness may reflect a generalised mammalian
preference for neotenic facial forms, but selection for neotenous faces
appears to have been enhanced in early Homo and was sustained during
the evolution of Middle and Upper Pleistocene members of our genus.
References
Adkins, R. & Jackson, R. 1978. Neolithic
Stone and Flint Axes from the River Thames. An Illustrated Corpus.
British Museum Occasional Paper No 1. London, British Museum.
Alley, T.R. & Cunningham, M.R. 1991. Averaged
faces are attractive, but very attractive faces are not average. Psychological
Science 2: 123-125.
Appleton, J. 1996. The Experience of Landscape.
Revised edition. New York, Wiley.
Balling, J.D. & Falk, J.H. 1982. Development
of visual preference for natural environments. Environment and Behavior
14: 5-28.
Behrensmeyer, A.K. 1978. The habitat of
Plio-Pleistocene hominids in East Africa. In Jolly, C. (ed.) Early Hominids
of Africa. London, Duckworth, pp. 165-189.
Burt, M. & Perrett, D.I. 1995. Perception of
age in adult Caucasian male faces. Computer graphic manipulation of shape
and colour information. Proceedings of the Royal Society of London.
B. 259: 137-143.
Clutton-Brock, J. 1981. Domesticated
Animals from Early Times. London, Heinemann.
Dissanayake, E. 1992. Homo Aestheticus.
Where Art Comes From and Why. New York, Free Press.
Enquist, M. & Arak, A. 1994. Symmetry, beauty
and evolution. Nature 372: 169-172.
Foley, R.A. 1996. An evolutionary and chronological
framework for human social behaviour. Proceedings of the British Academy
88: 95-117.
Jablonski, N.G. & Chaplin, G. 1993. Origin
of habitual terrestrial bipedalism in the ancestor of the Hominidae. Journal
of Human Evolution 24: 259-280.
Johnstone, R.A. 1994. Female preference for
symmetrical males as a by-product of selection for mate recognition. Nature
372: 172-175.
Jones, D. 1996. Physical Attractiveness and
the Theory of Sexual Selection. Ann Arbor, University of Michigan Museum
of Anthropology.
Jones, D., Hill, D. & Hill, K. 1993. Criteria
of facial attractiveness in five populations. Human Nature 4: 271-296.
Kaplan, S. 1992. Environmental preference in
a knowledge-seeking, knowledge-using organism. In Barkow, J.H., Cosmides,
L. & Tooby, J. (eds) The Adapted Mind. Oxford, Oxford University
Press, pp. 581-598.
Kingdon, J.S. 1980. The role of visual signals
and face patterns in African forest monkeys (guenons) of the genus Cercopithecus.
Transactions of the Zoological Society of London 35: 425-475.
Kohn, M. & Mithen, S. 1999. Handaxes: products
of sexual selection? Antiquity 73: 518-526.
Langlois, J.H. & Roggman, L.A. 1990. Attractive
faces are only average. Psychological Science1: 115-121.
Langlois, J.H., Roggman, L.A., Casey, R.J.,
Ritter, J.M., Rieser-Danner, L.A. & Jenkins, V.Y. 1987. Infant preferences
for attractive faces: rudiments of a stereotype? Developmental Psychology
23: 363-369.
Leakey, M.D. 1971. Olduvai Gorge Volume 3.
Excavations in Beds I and II, 1960-1963. Cambridge, Cambridge University
Press.
Leakey, M.G., Feibel, C.S., McDougall, I., Ward,
C. & Walker, A. 1998. New specimens and confirmation of an early age
for Australopithecus anamensis. Nature 393: 62-66.
Lorentz, K. 1971. Part and parcel in animal
and human societies: a methodological discussion. In Martin, R. (ed.) Studies
in Animal and Human Behavior. Volume 2. Cambridge, Ma: Harvard
University Press, pp. 115-195.
Manning, J.T. & Chamberlain, A.T. 1993.
Fluctuating asymmetry, sexual selection and canine teeth in primates. Proceedings
of the Royal Society of London B 251: 83-87.
Manning, J.T. & Chamberlain, A.T. 1994.
Fluctuating asymmetry in gorilla canines: a sensitive indicator of environmental
stress. Proceedings of the Royal Society of London B 255: 189-193.
McCabe, V. 1988. Facial proportions, perceived
age, and caregiving. In Alley, T.R. (ed.) Social and Applied Aspects
of Perceiving Faces. Hillsdale, Earlbaum, pp. 89-95.
Mithen, S. 1997. Cognitive archaeology, evolutionary
psychology and cultural transmission, with particular reference to religious
ideas. In Barton, C.M. and Clark, G.A. (eds) Rediscovering Darwin: Evolutionary
Theory and Archeological Explanation. Archeological Papers of the American
Anthropological Association No. 7. Arlington, American Anthropological
Association, pp. 67-74.
Møller, A.P. 1990. Female choice selects
for male sexual tail ornaments in the monogamous swallow. Nature
332:640-642.
Orians, G.H. & Heerwagen, J.H. 1992. Evolved
responses to landscapes. In Barkow, J.H., Cosmides, L. & Tooby, J.
(eds) The Adapted Mind. Oxford, Oxford University Press, pp. 555-579.
Parsons, P.A. 1992. Fluctuating asymmetry: a
biological monitor of environmental and genomic stress. Heredity
68: 361-364.
Perrett, D.I., May, K.A. & Yoshikawa, S.
1994. Facial shape and judgements of female attractiveness. Nature
368: 239-242.
Pfeiffer, J.E. 1982. The Creative Explosion.
New York, Harper and Row.
Phelps, M.T. & Roberts, W.A. 1994. Memory
for pictures of upright and inverted primate faces in humans (Homo sapiens),
squirrel monkeys (Saimiri sciureus), and pigeons (Columba livia).
Journal of Comparative Psychology 108: 114-125.
Stringer, C.B. & Andrews, P. 1988. Genetic
and fossil evidence for the origin of modern humans. Science 239:
1263-1268.
Symons, D. 1979. The Evolution of Human Sexuality.
New York, Oxford University Press.
White, T.D. 1988. The comparative biology of "robust"
Australopithecus:
clues from context. In Grine, F.E. (ed.) Evolutionary History of the
"robust" Australopithecines. New York, Aldine de Gruyter, pp. 449-479.
White, T.D., Suwa, G. & Asfaw, B. 1994. Australopithecus
ramidus, a new species of early hominid from Aramis, Ethiopia. Nature
371: 306-312.
WoldeGabriel, G., White, T.D., Suwa, G.,
Renne, P., de Heinzelin, J., Hart, W.K. & Heiken, G. 1994. Ecological
and temporal placement of early Pliocene hominids at Aramis, Ethiopia.
Nature 371: 330-333.
Wynn, T. 1995. Handaxe enigmas. World Archaeology
27: 10-24.
Wynn, T. 2000. Symmetry and the evolution of the
modular linguistic mind. In Carruthers, P. & Chamberlain, A.T. (eds)
Evolution
and the Human Mind. Cambridge, Cambridge University Press (in press).
Zahavi, A. & Zahavi, A. 1997. The Handicap
Principle. New York, Oxford University Press.
Return to top
* An earlier version of this paper was presented at
a conference on the Evolution of Mind, organised by the Hang Seng Centre
for Cognitive Studies, University of Sheffield, in June 1998.
Andrew Chamberlain can be contacted at: Department of Archaeology &
Prehistory, University of Sheffield, Northgate House, West St, Sheffield
S1 4ET.
mailto:[email protected]
Copyright © A. Chamberlain 2000
Copyright © assemblage 2000